

The majority of the world’s albatrosses and large petrels breed on islands in the Southern Ocean. It is well known that they can travel hundreds or thousands of kilometres on foraging trips. Potentially, this gives them access to the Antarctic seasonal sea-ice zone (SIZ), the area covered by sea ice for some of the year. This is one of the most extensive and productive marine habitats on Earth but also one of the most dynamic. Antarctic sea ice reaches its maximum extent – around 20 million km2 in September, covering around 40% of the Southern Ocean, but recedes to a minimum of around 4 million km2 by February. The spring melt releases nutrients, stabilises the water column and coincides with greater sunlight, together causing in a large pulse of primary production. This is followed after a lag of weeks to months by a pulse of secondary production and potentially therefore food for seabirds.
Historically, studies of seabirds in the SIZ were ship-based, gathering observations at fine scales but over relatively small spatiotemporal extents. These studies were able to describe broad patterns of habitat use such as the different species assemblages associated with different sea-ice zones, but little was known about the breeding status or provenance of seabirds in the SIZ. In this study, we aimed to use the large amount of data amassed by tracking albatrosses and large petrels breeding at Bird Island, South Georgia over the past thirty years to determine if and how subantarctic breeding seabirds use the SIZ.
In total we analysed 2,497 foraging trips made by 1,289 birds of seven species. Given that large Procellariiformes are hypermobile and have very protracted breeding seasons, we first wanted to establish whether there is a general movement to higher latitudes as the summer progresses due to the seasonal effects of sunlight on primary production. For four species, white-chinned petrels, black-browed albatrosses, grey-headed albatrosses and wandering albatrosses, this was indeed the case. Foraging latitude varied sinusoidally over the breeding season but was out of phase with peak solar irradiance by 2-3 months, presumably due to the time required for primary production to transfer to the mesopelagic species that these species feed on. Such seasonal latitudinal shifts, sometimes referred to as green wave surfing, are ultimately thought to be responsible for many large-scale migrations between breeding and non-breeding areas. However, as far as we are aware, this is the first evidence of it occurring during the breeding season.
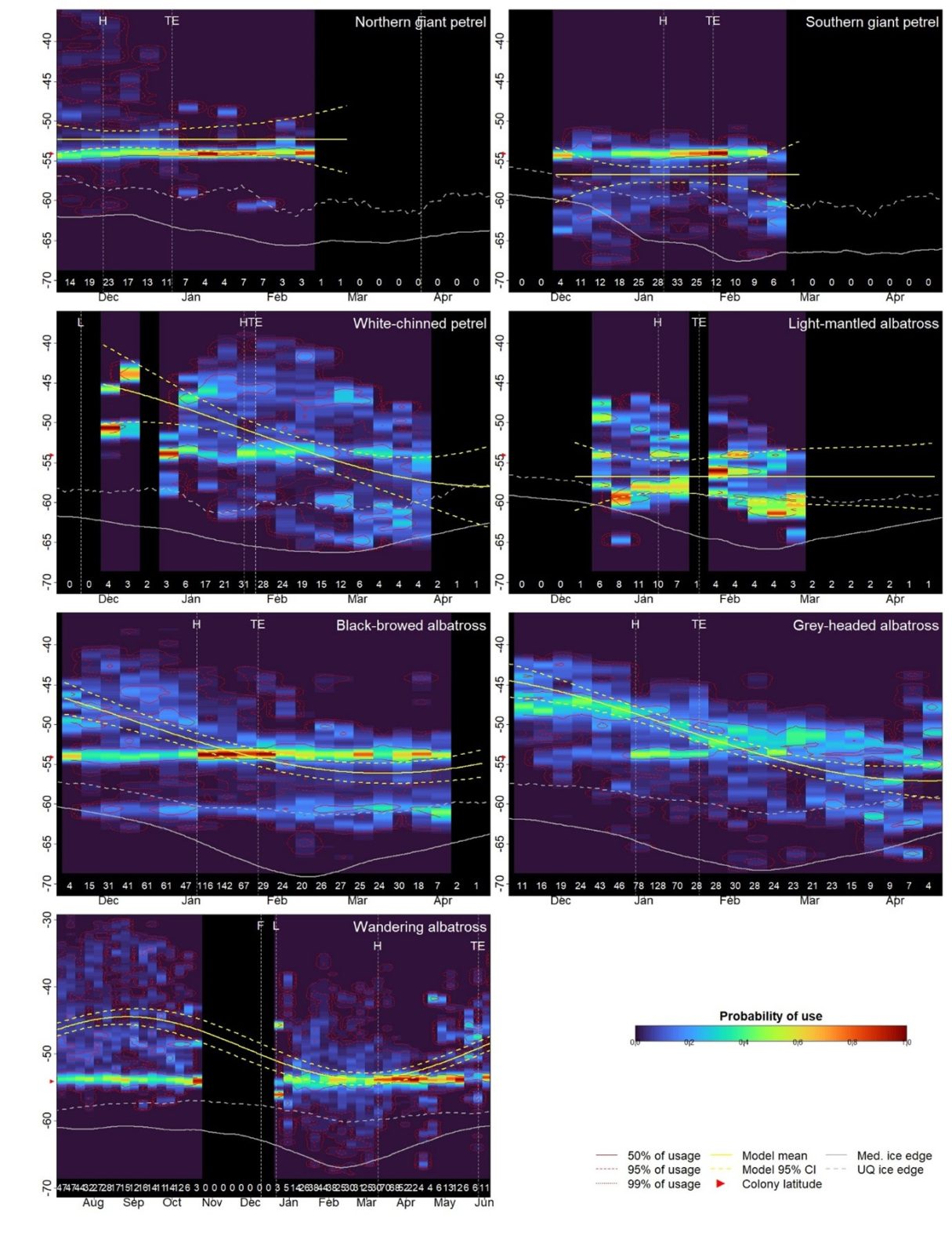
Figure 2 from Wakefield et al. 2024: Variation in foraging latitude over the breeding season. Time of year is on the x-axis and latitude is on the y-axis. Bird Island is located at 54° S. Colours show the probability of a latitude being used for foraging. Yellow lines show models of foraging latitude vs. time since midwinter and grey lines the median ice edge. L = laying, H = hatching, TE = thermal emancipation, F = fledging.
Foraging latitudes of northern and southern giant petrels and light-mantled albatrosses did not show these sinusoidal patterns, hinting that their movements may have other drivers. Next, we checked how much birds used sea-ice habitats and when they first used locations in the SIZ relative to when the spring melt occurred at those locations. Both giant petrel species used ice covered areas to some extent, especially in the spring. In particular, southern giant petrels used locations in the SIZ before the ice broke up there, sometimes travelling hundreds of km into the pack to areas with complete sea ice cover. We suspect that this was to scavenge seal pups or placentae because crabeater, Ross and leopard seals all give birth on ice floes between October and November. In contrast, light mantled albatrosses used parts of the SIZ only once they became ice-free, but they foraged right up to the ice edge. This species is often seen near icebergs so we speculate that they may specialise in foraging near floating ice, which often harbours communities of algae, crustaceans, fish, etc. on its undersides.
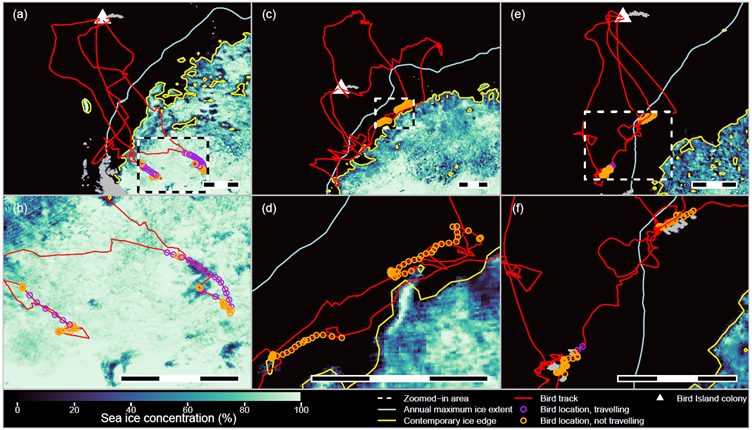
Figure 8 from Wakefield et al. 2024: Typical foraging trips made by albatrosses and large petrels in the Seasonal Sea-ice Zone. (a,b) southern giant petrels, (c,d) light-mantled albatrosses and (e,f) black-browed albatrosses. Bottom panels are zoomed in to the areas shown by the dashed square in the corresponding top panels.
The other albatross species and white-chinned petrels also used the SIZ but only after a lag of around six or seven weeks. This suggests two things: First, sea ice could act as a barrier to foraging or locomotion for these species. For example, the suppression of ocean waves could impede the dynamic soaring of large albatrosses, making them insufficiently manoeuvrable to land or take off between ice floes. Secondly, the lag suggests that time is required for the post-melt phytoplankton bloom to transfer to the trophic levels at which these species feed. Despite these limiting effects of sea ice itself, we found that the post-melt SIZ is an important foraging habitat for all the species we considered, including those such as wandering albatrosses that are not usually considered to be affected by sea ice. Indeed, we hypothesise that despite these species breeding hundreds of km from the SIZ, their phenology adapted to allow exploitation of this food-rich habitat during critical breeding stages.
Sea ice around Antarctica has remained relatively stable over satellite era, despite ongoing climate change. This is paradoxical, given the alarming declines in Arctic sea ice over the same period. Unfortunately, there are recent signs that Antarctic sea ice has also begun to decline, reaching record lows in each of the past three summers. Our findings are therefore timely, and we hope that they will stimulate further research into the linkages between Antarctic sea ice dynamics and the demography of subantarctic seabirds, especially as many of these already face unsustainable threats from bycatch.
Acknowledgements
This study was led by Durham University and the British Antarctic Survey in partnership with BirdLife International, the University of Barcelona, the University of Helsinki, Stony Brook University and the University of Coimbra. It was funded by the Leverhulme Trust, the European Research Council H2020, the Natural Environment Research Council (NERC), Darwin Plus, the National Science Foundation and the GSGSSI. It benefited from the strategic programme of the Marine and Environmental Sciences Centre (MARE), financed by the Foundation for Science and Technology (FCT) and represents a contribution to the Ecosystems component of the BAS Polar Science for Planet Earth Programme, funded by NERC.